In this essay we will discuss about the tissue systems in plants. After reading this essay you will learn about:- 1. Mechanical Tissue System in Plants 2. Vascular Tissue System in Plants 3. Epidermal Tissue System in Plants.
Essay # 1. Mechanical Tissue System in Plants:
Meaning of Mechanical Tissue System:
The tissue system that gives mechanical supports to the whole plant and their growing organs against different external and internal forces is called mechanical tissue system.
It is variously termed as stereome by Haberlandt (1918) and stereids by Schwendener (1874). Plant organs are subjected to various strains and stresses like bending and shearing stresses, stretching due to presence of large fruits, bending due to natural calamities like storm, heavy snow etc.
The stem has to withstand compression due to heavy weight of the large number of branches and leaves in the canopy. The branches, again have to withstand bending as they are oriented obliquely or horizontally. The fruit stalks tend to be extended due to weight of fruits and the roots are also subjected to extension when the stem bends due to strong wind.
Cell walls of all types of cells provide mechanical strength and rigidity to the plant. The woody plants achieve the structural stability and strength by the cell walls which contribute 95% of the dry weight of the wood.
The mechanical strength of non-lignified walls is due to the orientation of cellulose microfibrils. It is the major component of papeo cotton etc. Lignification in the walls of wood gives further strength. When cellulose is absent other wall polysaccharides may form microfibrils to give strength.
The mechanical strength is more in the direction parallel to the microfibrils in both types of cell walls. Due to secondary growth several layers of different microfibrillar orientation are developed. Therefore, the cell wall is strong enough to resist forces from any direction.
In stem the microfibrils of the thin-walled parenchymatous cells remain oriented transversely on the vertical walls so as to bend without break. In roots the microfibrils shows helical orientation to resist extensions.

Collenchyma and sclerenchyma cells, however, give maximum mechanical strength. Collenchyma walls get thickened at the corners or at the tangential walls due to the deposition of pectin, cellulose, hemicellulose, and protein.
As the collenchyma cells are living they can regulate the deposition and orientation of wall materials according to the needs of the developing organs. The collenchyma cells are also elastic due to the presence of hydrated pectin on the walls.

In some plants the branches and stems show special adaptation to resist the gravitational force by the formation of reaction wood. It differs in structure and location from ordinary wood. In conifers, this wood is located at the lower side of branches and stems to resist compression while in dicots it is present on the upper side of the branches to resist tension.
Principles Governing the Construction of Mechanical Tissue Systems:
The principle governing the construction of mechanical tissue system is the orientation of the mechanical tissues to obtain the maximum mechanical rigidity and elasticity with minimum expenditure of materials. This principle is observed in roots, stems, fruit stalks, branches etc.
Mechanical cells are developed to resist different strains and stresses. The resistances are designated as inflexibility (resistance to lateral bending), inextensibility (resistance to stretching), incompressibility (resistance to compression) and shearing stress (resistance to shearing action).
The principles of the construction and distribution of the mechanical tissues to form a system are:
1. Inflexibility:
Both the terrestrial arboreal plants as well as the herbs are encountered by forces to bend laterally. The plants with arborial habit resist such forces by the formation of heart- wood at the centre whereas the herbaceous plants resist through the formation of mechanical cells All the aerial organs are the inflexible organs as they face lateral bending strain due to application of lateral forces like high wind.
For inflexibility of plants the mechanical cells remain distributed in different organs like ‘I’-girders as railway lines, and also that are used in the construction of bridges, buildings etc. The girder consists of two upper and lower horizontal plates again connected by a vertical plate (Fig. 5.69).
In cross-sectional view it resembles the English capital letter T. The horizontal plates are called flanges and the connecting vertical plate is called the web. If a load is applied at the middle of I-girder a bending strain is encountered.
The upper flange tends to be compressed and the lower one tends to be extended. The flanges only suffer and resist the strains, the web on the other hand neither subjected to compression nor extension strains. This vertical line is termed as null-line as it is neutral regarding the strains.
A single I-girder can resist the lateral force in a single plane from both directions only. To resist such lateral forces from different directions several I-girders remain oriented in a circle perpendicular to the long axis. I-girders are arranged in such a way that their null-lines intercept at a common point forming a composite I-girder that can resist lateral forces from all directions.
This simple I-girder formation is the characteristic of many plants and thus they resist bending forces either singly or in suitable combinations. This I-girder formation is found in fibrovascular bundles where bands of sclerenchyma are present above and below the actual vascular strand.
These sclerenchymatous bands represent the upper and lower flanges and the vascular strand as the web. The fibrovascular bundles are present in the monocot leaves, e.g., Cyperus, where they are arranged in parallel. In the monocot stem, the vascular bundles are completely encircled by sclerenchyma termed bundle sheath.
In the cylindrical stems and other organs the mechanical cells remain at the periphery. In the hypodermis, collenchyma and sclerenchyma cells are found in dicot and monocot stems respectively. They are present either continuously or as isolated patches forming the flanges of composite I-girder.
In some members of Compositae sclerenchymatous bundle caps are present. In case of rectangular or square stems (e g Leonurus, Leucas etc.) the mechanical collenchyma cells are present at the four corners to form the flanges and the pith acts as the web.
Thus, diagonally opposite I-girders are formed. In Nyctanthes inverted vascular bundles are present at the four corners to act as flanges. In Zea mays stem sclerenchymatous bundle sheath is present and the stele is atactostele.
2. Inextensibility:
The underground anchoring organs like rhizomes, roots, etc. have to face longitudinal tensions. The fruit stalks, the climbers and lianes also have to withstand the longitudinal tension, as they are to bear the weight of fruits and their own weight, respectively, when they hang over the supporting object.
Many rooted hydrophytes have to be longitudinally stretched due to water current. The mechanical tissues in the centre of the hydrophyte -stems (e.g., Potamogeton lanceolatus), in the fruit stalks (e.g., Cucurbita) and in the lianes (e.g., Dioscorea) help to resist longitudinal stretching.
The centrally located mechanical tissues to resist longitudinal tension can be compared with electrical cable v. here rigidity is obtained from the central axile strands made up of metallic wires. This principle is observed in roots as well as in other inextensible organs. In roots the stele is small in comparison to the other external tissues and xylem and sclerenchyma are condensed within the stele.

3. Incompressibility:
The stems are encountered by the longitudinal compression developed by the weight of the canopy. This situation can be compared with a heavy load on the top of a cylindrical axis where the axis is subjected to longitudinal compression. The metallic rods remain condensed at the centre of the axis like the effective aggregation of the mechanical tissues at the centre of the stems.
The subterranean and submerged organs of plants are encountered by radial compression or crushing pressure by the surrounding medium like the soil and water, respectively. Mechanical cells in these organs remain at the periphery to resist such radial compression.

The stilt roots of Zea mays and Pandanus the mechanical cells are distributed as such so that they can resist both compression and extension forces. The stilt roots on the wind blowing side have to withstand longitudinal extension and those present on the opposite side have to resist longitudinal compression. Thus the same root is periodically compressed and extended.
In these roots in addition to centrally aggregated mechanical cells which resist longitudinal extension, sclerenchyma cells are present in the periphery to resist compression. Zea mays roots have peripheral sheet of sclerenchyma in the cortex (Fig. 5.69G) whereas Pandanus has developed isolated patches of sclerenchyma in the periphery of the cortex.
4. Shearing Stress:
The flat organs like leaves are often encountered by violent shearing stresses due to strong wind or water current. Such force acts at right angles to the surface of the leaves causing laceration. Dicot leaves are mechanically more resistant against such stress.
The I-girders present in them are firmly held together by a large number of cross-ties in the form of vein network (Fig. 5.70C). In monocot leaves parallel I-girders formed by fibrovascular bundles are present (Fig. 5.70B). Sclerotic strands are also present at the hypodermal region and leaf margins and the leaf blades are usually cuticularised to withstand shearing stress.
Essay # 2. Vascular Tissue System in Plants:
Meaning of Vascular Tissue System:
The vascular tissue system consists of the complex tissues, xylem and phloem, which constitute discrete conducting strands called vascular bundles. Each bundle is the isolated unit of the longitudinal strands of conducting tissues consisting essentially of xylem and phloem, frequently with a sheath of thick walled cells or other interspersed cells.
These are usually primary in nature. In spite of the mechanical support the vascular bundles primarily function in conduction — xylem for conduction of water with dissolved mineral matters, and phloem for conduction of prepared food matters in solution.

Occurrence:
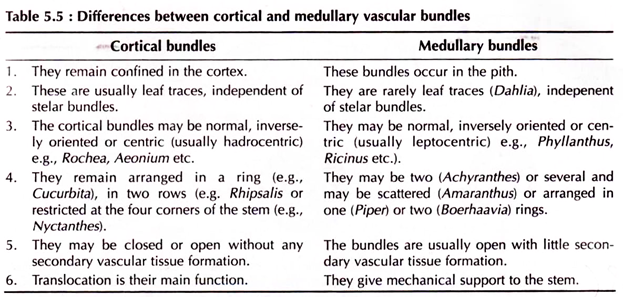
Every organ of the vascular plants possesses vascular bundles. In stems either they remain scattered throughout the ground tissue or may remain arranged in a compact ring in the stem axis to form the stele.
In some abnormal cases bundles may also be formed in the cortex or pith to form cortical (e.g., members of Cactaceae, Oleaceae) and medullary bundles (e.g., Dahlia, Piper etc.), respectively. In roots the xylems and phloems remain radially arranged at the centre.
Origin:
Vascular bundles originate from primary meristem. According to Hanstein and Schmidt plerome gives rise to the vascular bundles whereas the corpus gives rise to the nonvascular tissues. The procambial cells divide longitudinally and the peripheral derivatives form the phloem while the inner ones form the xylem (Haberlandt, 1914).
The first cells of the xylem initials mature into protoxylem whose development may be centripetal or centrifugal, or both, in relation to the centre of the axis of the organ and are termed as exarch, endarch and mesarch, respectively.
Exarch bundles are found in roots, endarch bundles are found in stems, while the mesarch bundles are found in stems of Tmesipteris and ferns. The xylem tissue formed later is termed as metaxylem. The first matured phloem is termed protophloem and it develops acropetally in relation to the centre of the axis. The lately formed phloem is termed metaphloem.
Types of Vascular Bundle:
According to the different types of arrangements of the xylem and phloem tissues, vascular bundles are of different types.
Following types of vascular bundes are found:
1. Collateral Bundle:
When xylem and phloem tissues remain on the same radius with reference to the centre point of the organ the phloem being external to the xylem — this type of vascular bundles are called collateral vascular bundle.
Again, depending on the presence or absence of the cambium, the vascular bundles are of the following types:
(a) Closed Collateral Bundle:
In this type of vascular bundles cambium is absent in-between xylem and phloem (Fig. 5.81) and, therefore, there is no further growth of the xylem and phloem tissues. Stems having this type of bundle do not have normal secondary growth as in monocotyledonous stems. Such closed vascular bundles usually remain enclosed within sclerenchymatous bundle sheath.
(b) Open Collateral Bundle:
When cambium is present in between the xylem and phloem tissues it is called open collateral vascular bundle (Fig. 5.81). This cambium is called fascicular cambium. The bundles can increase in diameter by normal secondary growth by the activity of the fascicular cambium. In dicotyledonous stem this type of vascular bundle is found.

2. Bicollateral Bundle:
If phloem tissue is situated on both the peripheral and central side of the xylem separated by strips of cambia in between is called bicollateral vascular bundle (Fig. 5.81).
So the sequence of vascular tissues in bicollateral bundles from the periphery is outer phloem, outer cambium, xylem, inner cambium and inner phloem. These bundles are obviously open type as two strips of cambia are present but the secondary growth is achieved only by the outer cambium as in Cucurbita stem.
3. Concentric Bundle:
When either of the vascular tissues surrounds the other or, vice versa, it is known as concentric bundle.
This type of vascular bundles are of the following types:
(a) Amphivasal Bundle:
In this type of bundle the xylem encircles the central strand of phloem and is also called leptocentric vascular bundle (Fig. 5.81). This type of bundle is found in Dracaena, Yucca etc.
(b) Amphicribral Bundle:
In this type the phloem encircles the central strand of xylem. It is also known as hadrocentric bundle (Fig. 5.81) as found in Selaginella. The concentric bundles either amphivasal or amphicribal are closed, as there is no cambium in between the two tissues.
4. Radial Vascular Bundle:
In this type of vascular bundle the primary xylem and phloem strands remain separated from each other by non-vascular tissues and are situated on alternate radii from the central point of the organ (Fig. 5.81). These bundles are the characteristic of roots.
There is no primary cambium in this type of bundle and the secondary thickening may occur by the secondary cambium originating during secondary growth in dicotyledonous root only. In dicot roots there are usually 4-6 arcs of xylem in contrast to monocot roots where there are more than six xylem arcs.



Essay # 3. Epidermal Tissue System in Plants:
Meaning and Origin of Epidermal Tissue System in Plants:
The covering cell layer(s) in the whole plant forms the epidermis which remains in direct contact with the environment. Therefore, this particular tissue adjusts itself with its surroundings because it has to protect the inner tissues from any adverse external factors like high temperature, desiccation, mechanical injury, pathogenic infections etc. The epidermis may persist throughout the plant life or it may, in some cases, be replaced by periderm when the older epidermis becomes inactive.
The outermost layer of apical meristem gives rise to the epidermis, which is variously named as protoderm, dermatogen and tunica by different authors. In cryptogams, however, it originates from solitary initial cell along with cortex and stele. The periblem gives rise to epidermis along with root cap in gymnospermous root, whereas in dicots the epidermis develops from dermatogen. The periblem in monocot roots develops the epidermis and the cortex.
Structure of Epidermal Tissue System in Plants:
Generally the epidermis is single layer of cells. Multiple-epidermis is the feature of Ficus and Nerium leaves and orchid root. The term velamen is given to the multiple epidermis of the orchid root. The multiple-epidermis is derived by the periclinal division of the epidermal initials.
Epidermal cell walls are usually thin, but thick-walled lignified epidermal cells occur in some gymnosperms. The outer surface of the epidermal cell wall may usually be cuticularised or waxy, giving rise to the characteristic surface features of the epidermis, which very often becomes a taxonomic characteristic.
The cuticle protects plants from desiccation as it is impervious to water. It also prevents the plant from various pathogens. Sometimes wax may be deposited on the surface of the cuticle to form a white powdery lining on various fruits (e.g., the plum, grapes etc.) and on leaves. Wax is deposited on the inside surface of the pitcher of Nepanthes in the form of overlapping scales to stick to the feet of the insects.
Epidermal cells are horizontally flattened and remain parallel to surface of the organ. In surface view the epidermal cell walls show sinuous outline in dicot leaves and straight or sinuous outline in monocot leaves. Usually the intercellular space is absent among the epidermal cells.
Chloroplastids are absent in epidermal cells except the specialised guard cells of stomata. Sometimes silicon may be deposited in the epidermal cells walls or in their lumens. Trichome cell walls may be silicified. Silicon is deposited in the bracts of rice, in the marginal trichomes of oat, in the leaves of Cyperus, Avena etc.
In Avena sativa the epidermal cells at the intercostals position of the internodes cork cells and silica containing cells remain together to form pairs. The epidermal cells may contain crystals of calcium carbonate to form cystolith (e.g., Ficus). These cells are called lithocysts.
Hairs:
Epidermal hairs are present in roots, stems, leaves, floral parts, seeds and stamens. Root hairs are present in the root hair zone behind the root tip of most monocots and dicots. They are formed in some species from distinct epidermal cells termed trichoblasts which protrude to form unicellular root hairs. The thin cell wall remains covered by a thin layer of cuticle. Mucilage may also occur on the surface.

The main function of root hairs is to absorb water and minerals in addition to anchorage. The root hairs synthesise cellulose at their tips.
Trichomes are the unicellular or multicellular hairy or glandular outgrowths of epidermal cells. Single linear row or several rows of cells constitute the multicellular trichome. The glandular ones are secretory in function.
The hairy trichomes give protection and also prevent water loss. The trichomes are also typified as stellate (star like) or dendroid (a miniature tree form, e.g., Verbascum). They may occur in tufts (e.g., Hamamelis). Hairy projections in flower petals are termed papillae.

Most trichomes possess thin and cellulosic or lignified cell walls. The glandular trichomes consist of unicellular or multicellular stalk and head with cuticle. Secretory substances accumulate beneath the cuticle (e.g., oils, resins, camphor, peppermint etc.) Sometimes scaly outgrowth with a short stalk and head consisting of discoid plate of cells may be formed (e.g., leaves of Olea). Some of the epidermal cells of Mesembryanthemum may enlarge where water accumulates. These specialised epidermal cells are termed as vesicles or bladders.
Bulliform Cells:
It is a group of outer larger epidermal cells found in monocot leaves easily distinguishable from other epidermal cells by their fan-shaped appearance in cross-section. The median cell of the group is the most elongated and the size of other cells on both side are gradually reduced (Fig. 5.73).
Individual cell of the group is thin-walled, hyaline and possesses a large vacuole but are devoid of chloroplastids. The cell walls are composed of cellulose and pectic substances. The outermost wall is cutinised and covered by cuticle. Bulliform cells are present in the outer epidermis of the leaves of Poaceae and other monocotyledons except Helobiae.

The suggested functions of bulliform cells are:
(i) The bulliform cells help to unroll the leaf during development,
(ii) Periodical turgour development in the bulliform cells causes the opening and closing of mature leaves,
(iii) These cells act as water reservoir, and
(iv) Due to silica deposition inside the bulliform cells and cuticularisation of their outer walls the leaves become mechanically rigid.